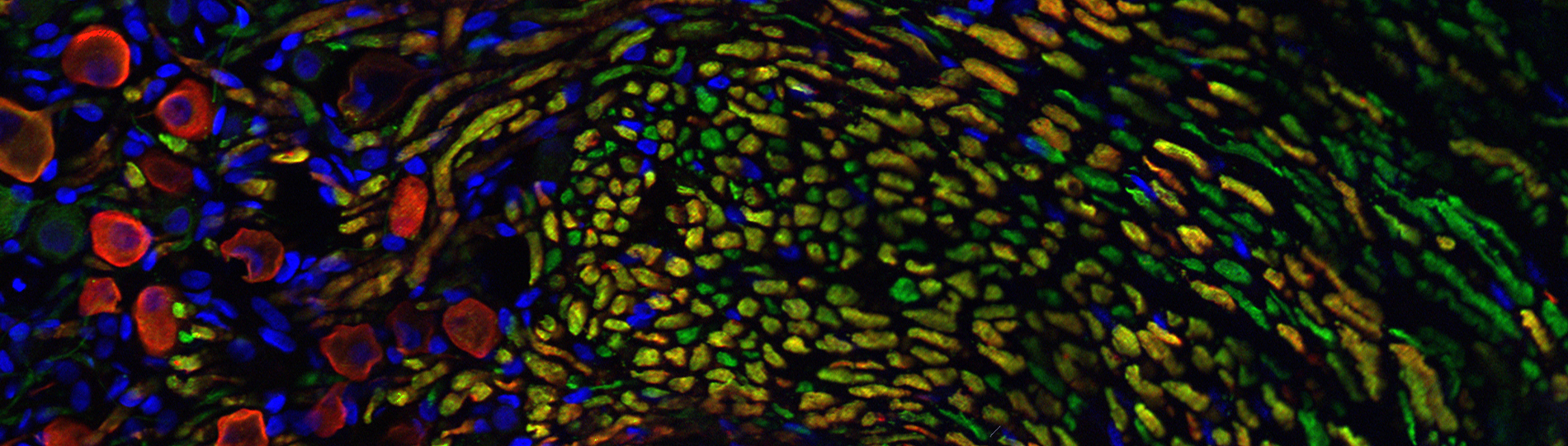
Synaptic Pharmacology of the Vestibular Apparatus
Many sensory systems are endowed with efferent feedback mechanisms that can modulate their primary input to the brain. That is, incoming information from a peripheral detector is delivered to a way station within the CNS whose output then modifies subsequent information coming from that same detector. Everyday examples include the pupillary reflex to bright light entering the eyes, the contraction of middle ear muscles to loud sounds, or the recruitment of additional muscle fibers when first lifting a heavy object. Here, the function of these efferent loops is presumably to optimize or tune each sensory modality to its stimulus. Additionally, these efferent pathways may also provide the appropriate circuitry for interactions with other sensory systems.
Sensory information regarding the position and movement of the head are encoded by the vestibular system, which begins as a number of small detectors called hair cells located within the inner ear. Like the preceding examples, the peripheral vestibular system is also endowed with a prominent efferent innervation. The functional role of this feedback system, however, is relatively unknown. We do know that when these efferent pathways are electrically stimulated, afferent output from vestibular endorgans is profoundly inhibited or excited, suggesting that vestibular efferents may be involved in both negative and positive feedback. If such efferent activity occurs under physiological conditions, it is almost certain to modify and transform vestibular information traveling to the CNS. Yet, very little information is available as to how and when these efferent actions ultimately impact the processing of vestibular information in an alert animal. Taking a reductionistic approach, this lab is addressing the function of the vestibular efferent system from four vantage points:
- Identifying the receptor mechanisms by which different efferent responses are generated during activation of their pathways;
- Characterizing how these efferent receptor mechanisms modulate afferent response properties by pairing afferent recordings during vestibular stimulation with activation of efferent pathways;
- Identification of efferent discharge patterns with direct, in vivo recordings from vestibular efferent neurons.
- Development of behavioral assays for monitoring and evaluating vestibular efferent function in alert animal models.
Individuals working in the lab can expect to learn neurophysiological, pharmacological, and immunohistochemical methods for studying vestibular synaptic transmission in several animal models, and the necessary computational techniques for analyzing these data.

J. Christopher Holt, Ph.D.
Principal Investigator


- Elucidating the Role of Muscarinic Acetylcholine Receptor Signalling in Efferent-Mediated Responses of Mammalian Vestibular Afferents.; The European journal of neuroscience; Vol 63(10), pp. e70540. 2026 May.
- Persistence of vestibular function in the absence of glutamatergic transmission from hair cells.; Scientific reports. 2026 Mar 23.
- Systemic Calcitonin Gene-Related Peptide (CGRP) modifies auditory and vestibular end organ electrical potentials, and increases sensory hypersensitivities.; Journal of neurophysiology. 2025 May 27.
- Outer hair cells stir cochlear fluids.; eLife; Vol 13. 2025 Jan 16.
Affiliations
- Otolaryngology
- Neuroscience
- Del Monte Institute for Neuroscience
- Cellular and Molecular Pharmacology and Physiology Ph.D. Program
- Master’s Degree in Pharmacology
- Neurobiology & Anatomy Ph.D. Program
- Neuroscience Ph.D. Program
- PhD Program in Pathology - Cell Biology of Disease
- Hearing and Balance Research Collaborative
Contact Us
Holt Lab
MC 6-7556
601 Elmwood Ave
Rochester, NY 14642
(585) 276-5334