CGRP Biosynthesis
Most neuropeptides are synthesized as large, biologically inactive precursors that must undergo a series of post-translational modifications to produce the smaller biologically active peptides. Modifications include endoproteolysis at pairs of basic amino acids, removal of the basic residues that constitute the cleavage site, carboxyl amidation and amino acetylation and cyclization. Endoproteolysis can exhibit tissue and developmental specificity, and is a key step in regulating peptide hormone secretion. Work in my laboratory has focused on determining the substrate-specificity of endoproteolysis, and on identifying the neuroendocrine endoproteases responsible for cleavage.
We have studied the specificity of propeptide endoproteolysis by introducing mutations into propeptide cDNAs which alter their basic amino acid cleavage sites, and analyzed the ability of the mutated propeptides to be cleaved in transfected neuroendocrine cell lines. Cleavage at tetrabasic sites is an important regulatory step in the processing of proopiomelanocortin (POMC) in the pituitary, where tissue-specific cleavage at the LysLysArgArg site in POMC produces either adrenocorticotropin (ACTH) in the anterior pituitary, or α-melanocyte stimulating hormone (αMSH) in the intermediate pituitary. We have used procalcitonin gene-related peptide (proCGRP) as our model system for these studies (Fig. 1). Following removal of a 25 aa signal peptide, the 103 aa proCGRP molecule is cleaved at two sites, a dibasic site (Lys56Arg57) and a tetrabasic site (Arg96Arg97Arg98Arg99) to produce a 37 aa carboxyl-amidated neuropeptide, CGRP.
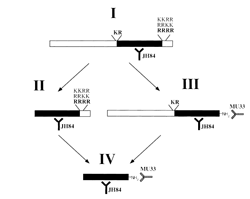
Figure 1. proCGRP is cleaved at two sites and
carboxyl-amidated to produce mature bioactive peptide.
Two antibodies used for study recognized CGRP
and proCGRP (JH84) or carboxyl-amidated CGRP (MU33).
We used the Polymerase Chain Reaction (PCR) was used to introduce mutations into the proCGRP cDNA which encoded alterations in the tetrabasic cleavage site, and the mutated cDNAs were cloned into the eukaryotic expression plasmid pCMV.Neo. AtT-20 cells (derived from mouse anterior pituitary corticotroph cells) and GH3 cells (derived from rat anterior pituitary somatomamotroph cells) were transfected, and stable cell lines were obtained following selection with the antibiotic G-418 (geneticin). Cells were then biosynthetically labeled with 3H-Leu, and proCGRP-derived peptides were immunoprecipitated with antibodies raised against the CGRP peptide, and the immunoprecipitates were analyzed by SDS-PAGE and HPLC. AtT20 express POMC, but do not cleave the LysLysArgArg site, similar to the anterior pituitary. Interestingly, AtT20 cells did cleave mutated proCGRP at this site. Cleavage of LysLysArgArg in exogenous proCGRP, but not in endogenous POMC, suggests that the specificity of cleavage at tetrabasic sites is not defined solely by the endoproteases expressed by the cell or by the amino acid sequence at the cleavage site, but is also dependent on the structure of the propeptide.
Determination of the tissue distribution and developmental regulation of the prohormone convertases responsible for maturation of propeptides has been an arduous task because of the complexity of most vertebrates species. To overcome this limitation, an alternative approach is to study the regulation of these endoproteases in a simpler, well characterized organism. Such an organism is the free-living nematode Caenorhabditis elegans, which has many genetic mutants described, strains available in culture, complete cell lineage mapped from the egg to the adult, and complete anatomy described at the ultrastructural level. However, very little is known about serine endoproteases in C. elegans.
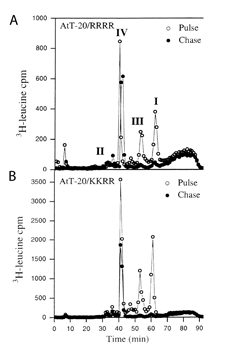
Figure 2. A. Tetrabasic site LysLysArgArg is efficiently
cleaved in transfected AtT20 cells expressing mutated proCGRP;
B. Pulse-chase experiment shows that propeptide (peak I) is
converted to mature CGRP (peak IV) as efficiently as is proCGRP
containing wild-type ArgArgArgArg site.
The bli-4 gene encodes a protein which is involved in cuticle development and is homologous to Kex2-like endoproteases. Therefore, we developed a cloning strategy based on a combination of reverse transcription-polymerase chain reaction (RT-PCR) and rapid amplification of cDNA ends (RACE) to clone several of the Kex2-like endoproteases involved in neuropeptide biosynthesis. To facilitate developmental studies on endoprotease expression, we have cloned prohormone convertase 2 (PC2) and furin from the nematode Caenorhabditis elegans. These studies on neuropeptide biosynthesis have determined the molecular basis for tissue-specific synthesis of neuropeptide hormones in the pituitary, and delineated the neuroendocrine endoproteases in C. elegans, an organism ideal for future studies on the developmental regulation of prohormone convertases.