Mac-1 (CD11b/CD18) in Neutrophil Migration
Mac-1 (CD11b/CD18) in Neutrophil Migration
Neutrophils play key roles in the host defense network against pathogens by virtue of their abilities to phagocytose microorganisms and to produce oxygen free radicals and proteolytic enzymes. Extravasation of neutrophils from the blood stream proceeds through three coordinated steps: rolling and tethering, firm adhesion, and transmigration. The first step depends on the selectin molecules expressed on both neutrophils and endothelial cells (EC). The second step is mediated through interactions of the β2 integrins, including LFA-1 and Mac-1, with their counter receptors, ICAM-1 and ICAM-2, on the EC. Then the process is followed by an additional step, called “transendothelial migration”, by which neutrophils leave the blood stream and enter a site of inflammation. The importance of the β2 integrin family in vivo is illustrated by patients with inherited leukocyte adhesion deficiency (LAD), who have a genetic defect in β2 integrin expression. LAD patients have recurrent, life-threatening bacterial infections and impaired wound healing and are candidates for bone marrow transplantation early in life.
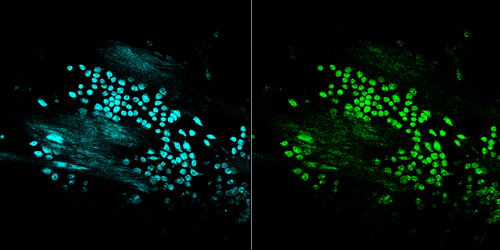
CFP and YFP images of cells in the Cremaster venule of live mouse.
Recent studies of leukocyte trafficking have generated substantial insights into the physiologic and pathologic migration patterns of leukocytes and their role in health and disease. This information has been essential for the development of new anti-inflammatory therapies, such as monoclonal antibodies against leukocyte integrins to treat psoriasis and multiple sclerosis. Among the established techniques to study leukocyte trafficking, direct observation of leukocyte rolling and migration by intravital microscopy (IVM) is one of the most important experimental approaches. One method for studying distinct leukocyte subsets consists of purifying cells from donor mice, labeling them fluorescently in vitro and re-injecting them into a recipient. Transgenic mice expressing green fluorescent protein (GFP) or its variants in a cell type-specific fashion are alternatives to ex vivo purified, fluorescently labeled cells.
This approach subjects cells to less ex vivo handling and genetically encoded fluorescent tags do not become diluted or fade in dividing cells. Additionally, fluorescent protein transgenic leukocytes can be studied endogenously without ex vivo manipulation or adoptive transfer. In vivo fluorescent sensors that are based on FRET hold promise for IVM to study cell activation and/or signaling during long-term cell migrations. We are currently generating variety of fluorescence protein knock-in mice for studying dynamic neutrophil migration and integrin activation in live animals.