URMC / Labs / Holt Lab / Projects / Characterizing-the-morphophysiological-properties-of-vestibular-efferent-neurons
Characterizing the Morphophysiological Properties of Vestibular Efferent Neurons
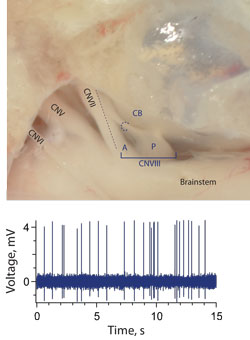
Anatomical location for direct recordings from efferent neurons. In our animal
model, cranial nerve VIII splits into a posterior and anterior branch immediately after
exiting the brainstem (Top panel). Most, if not all, efferent fibers travel in the anterior
branch. Shortly before entering the otic capsule, those efferent fibers destined for the
posterior portions of the inner ear cross over and join the posterior branch. This
crossing branch is referred to as the crossbridge (CB). The CB provides a convenient
and relatively accessible location to record efferent activity. Activity recorded from a
presumed efferent neuron in the CB is shown in the lower panel.
In order to define a functional role for vestibular efferents, it is essential to know what their discharge properties are. This certainly includes descriptions of any spontaneous
or background activity, discharge regularity, and what the upward limits of their discharge might be. It also becomes important to determine if we can modify efferent discharge by modulating afferent activity. Vestibular efferents may exhibit background activity which responds to both vestibular and extralabyrinthine sensory inputs. Such activity, by recruiting efferent receptors on hair cells and afferent fibers, would, in turn, modify afferent physiology. Our working hypothesis is that efferent neurons are normally active and contribute to the resting discharge and sensitivity of afferent fibers. To test our hypothesis, recordings will be made from both efferent and afferent fibers at rest and during stimulation of sensory input from both vestibular afferents and cranial nerves.
In our working model, vestibular efferent fibers are anatomically isolated within a structure called the cross-bridge where single unit activity can be measured with sharp microelectrodes (see Figure). Here, we can record from the cross-bridge and identify efferents by antidromically stimulating vestibular nerve branches. We also plan to test if standing efferent discharge influences afferent sensitivity or discharge by recording from posterior canal afferents before and during the application of anesthetic agents and cholinergic antagonists know to block efferent function. Because of the possibility that efferents innervating different regions of the crista may differ in their discharge properties, recorded efferent neurons will be dye-filled to identify terminal morphology and innervation patterns.
« back to all projects